Understanding which species are going to establish within a given community (i.e. invasion) is one of the most fundamental pursuits in ecology. In today’s era of global change, this is especially important given the threat of exotic species to native biodiversity. Many hypotheses have been developed in attempts to answer this question, but two predominate. The first is that newly establishing species need to be similar enough to the species at the site to survive the environmental conditions at that site, but dissimilar enough to avoid strong competition with the resident species for the limited available resources (i.e. Darwin’s naturalization hypothesis). The second hypothesis is largely an extension of the first. It predicts that diverse plant communities should be more resistant to invasion because most of the available resources will already be in use due to the larger number of, presumably functionally distinct, species. However, there are flaws in these hypotheses that limit their applicability.
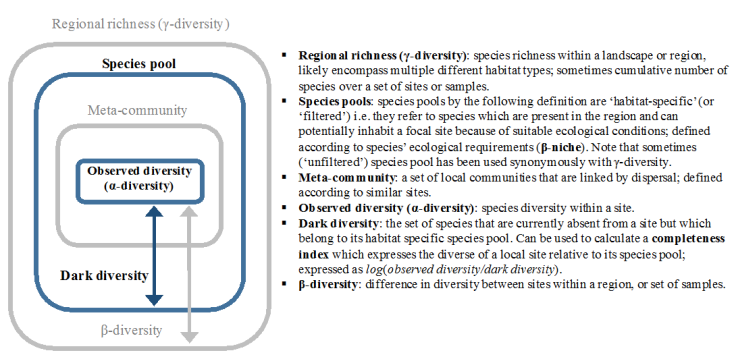
The first major flaw is that the species found at a given site are not entirely the product of interactions among those species: they are also influenced by the regional processes and environmental filtering which combine to shape the species pool for that site (Fig. 1). These processes also have their own effects on invasion. Sites that can support a greater number of species may also support a greater number of invaders (Fig. 1). Consequently, sites that are more diverse due to a larger species pool may also be more readily invaded, complicating any use of local diversity as an indicator of potential biotic resistance. However, if we make local richness proportional to the size of the species pool using the community completeness index (Fig. 1), we may find a more consistent relationship between diversity and invasion.
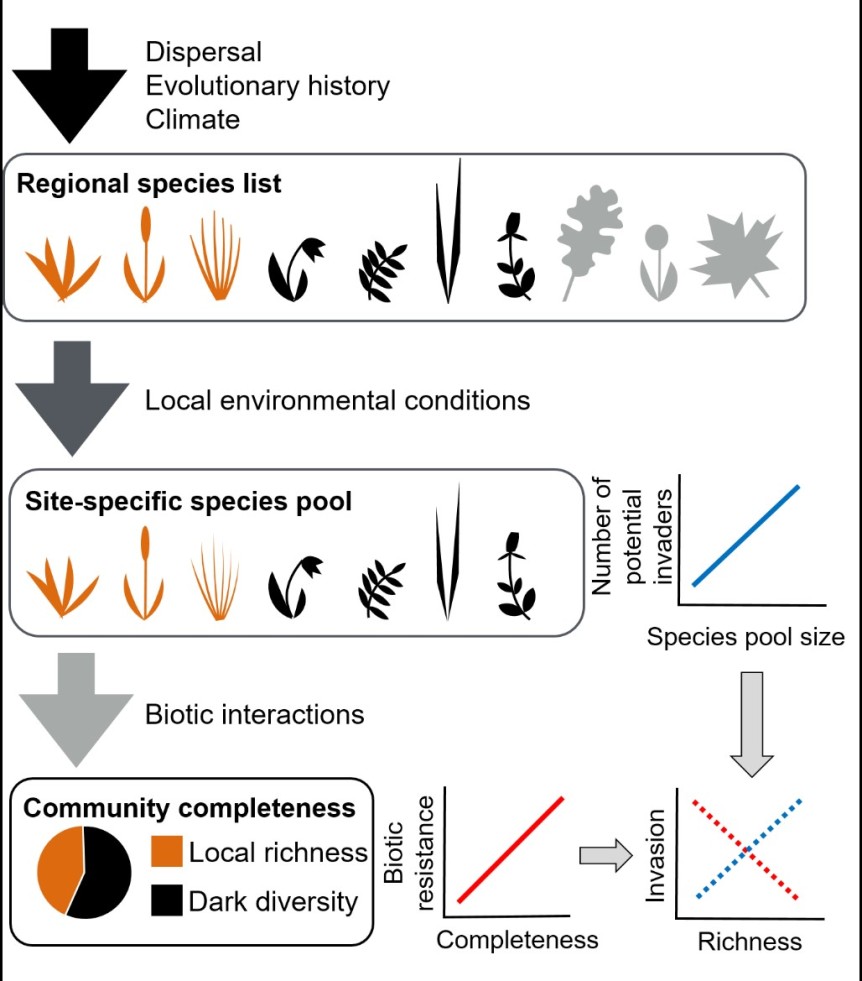
Using a large experiment, we tested this hypothesis by adding both native and exotic species to sites distributed throughout southern Estonia (Bennett et al. 2016 Ecology Letters). We found that by accounting for the size of the site-specific species pool and community completeness, we were better able to explain the variation in invasion than by using local richness alone. Interestingly, species pool size was not always a positive indicator of invasion rates. For exotic species, a larger species pool sometimes resulted in greater biotic resistance. A larger species pool may contain a larger number of strong competitors by random chance alone. However, if competition trades off with dispersal abilities, as it sometimes does, then species pools that remain small due to limited dispersal may be less likely to contain any species that can resist the invader.
The previous result highlights the importance of understanding the characteristics of the species in different species pools for predicting invasion. Darwin’s naturalization hypothesis attempts to do so, by comparing the characteristics of the potential invader to the resident community. However, this approach does not consider species pool effects on local diversity. Further, it is based on a flawed premise, as competition is not necessarily strongest between the most similar species. Instead competition will depend on the underlying assembly dynamics within the community. In a recent paper (Bennett and Pärtel 2017 Ecology and Evolution), we developed a new method that models environmental and biotic filtering by comparing species across multiple components of the regional species list (Fig. 2).
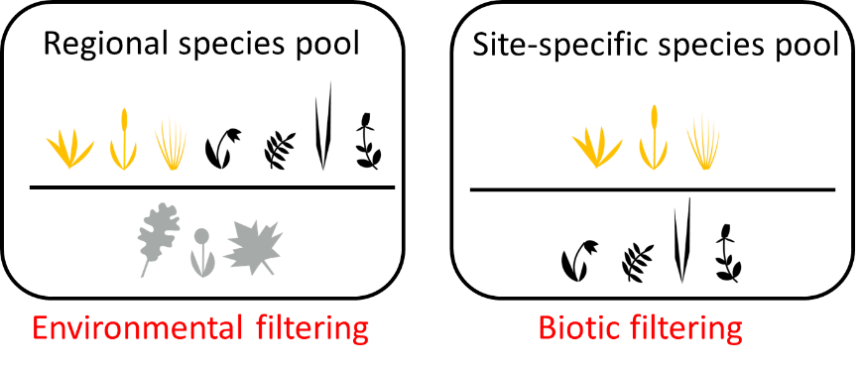
We measure the similarity among species within the site-specific pool relative to the similarity between the site-specific pool and the remainder of the regional species list to model environmental filtering. Similarly, we compare local diversity to the species from the site-specific species pool that are currently absent (dark diversity) to model biotic filtering. Using these models, and the similarity between the invader and these two components of diversity, we can then predict which invaders should establish (Fig. 3). When we tested the method using data from experiments that added seed to Estonian grasslands, we were able to predict approximately 50% of establishment from seed – a very impressive result. This result was based on only native species, but we are working to test the model using exotic species as well.
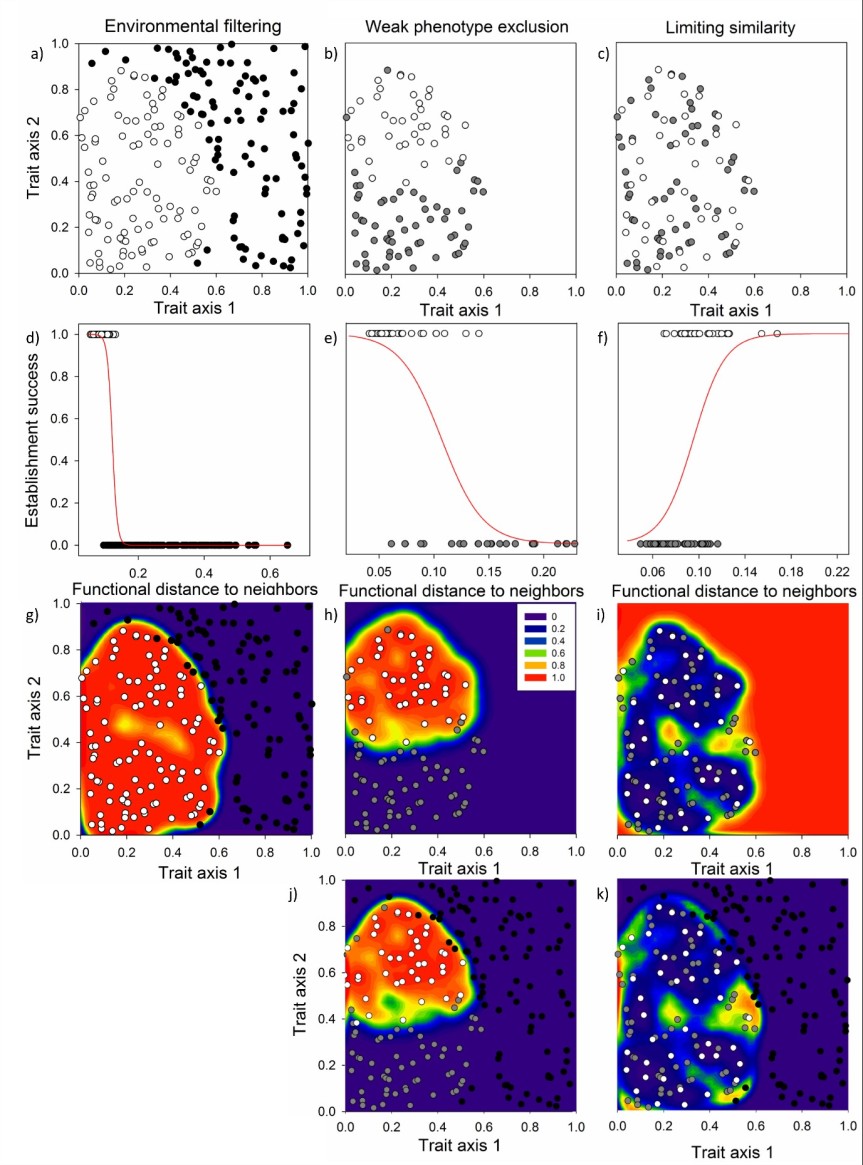
These two papers provide strong evidence that species pools need to be considered when trying to understand and predict invasion. Further, they also indicate a strong need to re-evaluate many of the hypotheses in invasion biology in the light of current knowledge on community assembly.
Bennett, J.A., Riibak, K., Kook, E., Reier, Ü., Tamme, R., Guillermo Bueno, C. & Pärtel, M. (2016) Species pools, community completeness and invasion: disentangling diversity effects on the establishment of native and alien species. Ecology Letters, 19, 1496-1505.
, . (2017) Predicting species establishment using absent species and functional neighborhoods. Ecology and Evolution, 7, 2223–2237.